¶ Substantia Nigra
¶ Introduction
Substantia Nigra is an important component in the neurobiology of neurodegenerative diseases. This page provides detailed information about its structure, function, and role in disease processes. [@zecca2006]
¶ Overview
The substantia nigra (Latin for "black substance") is a midbrain structure that plays a critical role in motor control, reward processing, and cognitive function. Its distinctive dark appearance results from high concentrations of neuromelanin, a pigment that accumulates in dopaminergic neurons with age (Zecca et al., 2003. The substantia nigra is one of the most prominently affected brain regions in parkinsons, where progressive loss of its dopaminergic-neurons-snpc leads to the characteristic motor symptoms of the disorder (Damier et al., 1999. [@damier1999]
¶ Anatomy and Organization
¶ Location and Gross Structure
The substantia nigra is located in the ventral midbrain, dorsal to the cerebral peduncles and ventral to the red-nucleus-expanded (Lanciego et al., 2012. It is a paired structure, with one substantia nigra on each side of the midline. The substantia nigra extends from the upper brainstem through the midbrain and is part of the basal-ganglia circuit, though it lies anatomically outside the basal ganglia proper. [@lanciego2012]
¶ Pars Compacta (SNc)
The dorsal tier of the substantia nigra, the pars compacta, contains densely packed, neuromelanin-pigmented dopaminergic-neurons-snpc that are the principal source of dopamine for the dorsal striatum (caudate-nucleus and putamen) (Björklund & Dunnett, 2007. The SNc is estimated to contain approximately 400,000–600,000 dopaminergic neurons in each hemisphere in healthy individuals (Pakkenberg et al., 1991. These neurons project through the nigrostriatal pathway — the mesostriatal dopaminergic system — which is essential for the initiation and modulation of voluntary movement. [@bjrklund2007]
SNc dopaminergic neurons are characterized by: [@pakkenberg1991]
- Expression of tyrosine hydroxylase (TH), the rate-limiting enzyme in dopamine synthesis
- Accumulation of neuromelanin pigment (visible as the dark band on gross examination)
- Extensive axonal arbors — a single SNc neuron can form 100,000–250,000 synaptic connections in the striatum (Matsuda et al., 2009
- Autonomous pacemaker activity, driven by L-type calcium channels
¶ Pars Reticulata (SNr)
The ventral tier, the pars reticulata, contains primarily GABAergic neurons that constitute one of the two major output nuclei of the basal-ganglia (the other being the globus-pallidus interna) (Deniau et al., 2007. SNr neurons are tonically active, firing at rates of 60–90 Hz, and exert continuous inhibition on their targets in the thalamus, superior-colliculus, and pedunculopontine-nucleus. [@matsuda2009]
The SNr receives GABAergic input from the striatum via the direct pathway (striatonigral projection). When striatal medium spiny neurons are activated by cortical input and dopaminergic facilitation, they inhibit the SNr, which in turn disinhibits thalamic neurons — allowing movement to proceed. Disruption of this circuit underlies the motor symptoms of parkinsons. [@deniau2007]
¶ Pars Lateralis
A third, less well-defined region — the pars lateralis — has been identified in some classifications. It contains a mixed population of neurons and is thought to contribute to non-motor (limbic and associative) functions (Halliday, 2004. [@gerfen2011]
¶ Neurochemistry and Dopamine Synthesis
¶ Dopamine Pathway
The substantia nigra pars compacta is the origin of the nigrostriatal dopamine pathway, which is the primary dopaminergic innervation of the dorsal striatum. dopamine synthesis proceeds through the following steps: [@zhang2011]
- L-tyrosine is transported into the neuron
- Tyrosine hydroxylase (TH) converts tyrosine to L-DOPA (the rate-limiting step)
- Aromatic L-amino acid decarboxylase (AADC) converts L-DOPA to dopamine
- Dopamine is packaged into synaptic vesicles by the vesicular monoamine transporter 2 (VMAT2)
Released dopamine acts on two main receptor families in the striatum: [@kalia2015]
- D1 receptors (direct pathway): excitatory effect on striatonigral neurons, facilitating movement
- D2 receptors (indirect pathway): inhibitory effect on striatopallidal neurons, suppressing unwanted movements
This dual-pathway model explains why dopamine depletion in parkinsons leads to both bradykinesia (reduced D1 stimulation) and rigidity (increased D2-pathway activity) (Gerfen & Surmeier, 2011. [@spillantini1997]
¶ Neuromelanin
Neuromelanin is a dark, insoluble pigment that accumulates in dopaminergic neurons of the SNc throughout life. It is formed from the oxidation of excess cytoplasmic dopamine and related catecholamines (Zecca et al., 2003. Neuromelanin has dual roles: [@braak2003]
- Neuroprotective: chelates iron and sequesters toxic metabolites
- Potentially neurotoxic: released neuromelanin from dying neurons activates microglia | Striatopallidal (indirect) | gaba |
| Subthalamic nucleus | Subthalamonigral | glutamate | [@surmeier2017]
| Cerebral [cortex | Corticonigral | glutamate | [@pacelli2015]
| Pedunculopontine nucleus | Tegmentonigral | acetylcholine / Glutamate | [@chan2007]
| Dorsal raphe nucleus | Raphenigral | serotonin | [@dexter1989]
¶ Efferent Projections
- SNc → striatum (nigrostriatal): Dopaminergic; the major output of the pars compacta
- SNr → thalamus: GABAergic; targets ventral anterior and ventrolateral nuclei
- SNr → Superior colliculus: GABAergic; controls saccadic eye movements
- SNr → Pedunculopontine nucleus: GABAergic; influences locomotion
¶ Role in Parkinson's Disease
¶ Neurodegeneration of SNc Dopaminergic Neurons
Parkinson's disease is pathologically defined by the progressive loss of dopaminergic neurons in the substantia nigra pars compacta. By the time motor symptoms appear, approximately 50–70% of SNc dopaminergic neurons have already been lost, and striatal dopamine levels are reduced by 60–80%[@kalia2015]. The loss follows a characteristic ventrolateral-to-dorsomedial gradient within the SNc, with the ventrolateral tier (projecting to the dorsal putamen) being most severely affected[@damier1999]. [@mcgeer1988]
¶ Lewy Body Pathology
Surviving SNc neurons in PD frequently contain Lewy bodies — intracellular inclusions composed primarily of misfolded alpha-synuclein[@spillantini1997]. Lewy bodies and Lewy neurites are pathological hallmarks of PD and are believed to represent a cellular response to protein-aggregation stress. The prion-like-spreading hypothesis proposes that pathological alpha-synuclein propagates through connected neural circuits in a stereotyped pattern, as described by Braak staging[@braak2003]. [@hirsch2009]
¶ Selective Vulnerability
The selective vulnerability of SNc dopaminergic neurons to degeneration in Parkinson's disease is a central question in the field[@surmeier2017]. Several factors contribute to the vulnerability of these neurons (see also Selective Neuronal Vulnerability)[@booth2017]:
- High metabolic demand: SNc neurons have extremely large axonal arbors and autonomous pacemaking activity, resulting in basal oxygen consumption rates 3-fold higher than ventral tegmental area (VTA) dopaminergic neurons[@pacelli2015]
- Calcium oscillations: Pacemaker activity relies on L-type Cav1.3 calcium channels, leading to sustained intracellular calcium influx and mitochondrial stress[@chan2007]
- Dopamine metabolism: Cytoplasmic dopamine generates oxidative stress through auto-oxidation and enzymatic metabolism
- Iron accumulation: The substantia nigra has the highest iron concentration of any brain region, and iron levels increase with age, promoting ferroptosis and oxidative damage[@dexter1989]
- Reduced trophic support: Decreased expression of GDNF and BDNF in the aging SNc
¶ Genetic Factors
Multiple genes implicated in familial Parkinson's disease converge on pathways active in SNc neurons[@sulzer2018]:
- SNCA (alpha-synuclein): Mutations or multiplications cause autosomal dominant PD with Lewy body pathology
- LRRK2 (LRRK2 kinase): Most common genetic cause of PD; regulates vesicle trafficking and autophagy
- PRKN (parkin) and PINK1 (PINK1 kinase): Autosomal recessive PD; critical for mitophagy and mitochondrial quality control
- PARK7 (DJ-1): Oxidative stress sensor and mitochondrial protector
- GBA (glucocerebrosidase): Lysosomal enzyme; mutations are the most common genetic risk factor for PD
¶ neuroinflammation in the Substantia Nigra
¶ Microglial Activation
The substantia nigra shows prominent microglial/cell-types/microglia activation in [parkinsons, as first demonstrated by McGeer et al. (1988) through HLA-DR immunostaining (McGeer et al., 1988. Activated [microglia in the SNc release pro-inflammatory mediators including: [@limousin1998]
- Tumor necrosis factor-alpha (TNF-α)
- Interleukin-1 beta (IL-1β) and interleukin-6 (IL-6)
- Reactive oxygen and nitrogen species
- Prostaglandins
This chronic neuroinflammation creates a feed-forward cycle: neuronal death releases neuromelanin and aggregated alpha-synuclein, which further activate [microglia, amplifying the inflammatory response (Hirsch & Hunot, 2009. [@kamath2022]
¶ Astrocyte Dysfunction
astrocytes]astrocytes] in the substantia nigra undergo reactive changes in Parkinson's Disease, with increased glial-fibrillary-acidic-protein (glial-fibrillary-acidic-protein expression (Booth et al., 2017. Astrocytic dysfunction impairs: [@agarwal2024]
- Glutamate buffering, contributing to excitotoxic stress
- Potassium homeostasis
- Metabolic support to dopaminergic neurons
- blood-brain-barrier integrity
¶ Other Diseases Affecting the Substantia Nigra
Beyond parkinsons, the substantia nigra is affected in several other conditions:
- lewy-body-dementia: SNc dopaminergic loss plus widespread cortical Lewy body pathology
- msa: SNc degeneration with glial cytoplasmic inclusions in oligodendrocytes
- Progressive Supranuclear Palsy: Neuronal loss in the SNc with tau] pathology
- corticobasal-degeneration: Variable SNc involvement with asymmetric tau] deposition
- neurodegeneration-brain-iron-accumulation: Iron overload in the substantia nigra and globus pallidus
¶ Therapeutic Approaches Targeting the Substantia Nigra
¶ Pharmacological
- levodopa: The gold standard for PD treatment; a dopamine precursor converted to dopamine in remaining SNc neurons and striatal terminals
- dopamine-agonists: Directly stimulate dopamine receptors in the striatum, bypassing the need for dopaminergic neurons
- MAO-B inhibitors: Reduce dopamine degradation, prolonging its action (see [MAO-B Inhibitors/therapeutics/mao-b
¶ Surgical and Neurostimulation
- deep-brain-stimulation: High-frequency stimulation of the subthalamic-nucleus or globus pallidus interna modulates basal ganglia output, alleviating motor symptoms (Limousin et al., 1998
- focused-ultrasound: Emerging non-invasive approach for targeted neuromodulation
¶ Neuroprotective and Restorative Strategies
- Neurotrophic factor delivery: gdnf and neurturin infusion to promote SNc neuron survival
- Cell replacement therapy: Transplantation of stem cell-derived dopaminergic neurons into the striatum (see stem-cell-therapy
- gene-therapy: AAV-mediated delivery of AADC, GDNF, or alpha-synuclein-targeting constructs
- alpha-synuclein immunotherapy: Antibodies targeting aggregated alpha-synuclein (see immunotherapy
¶ Detailed Anatomy and Cellular Organization
¶ Pars Compacta Subdivisions
The SNc is not a homogeneous structure but comprises molecularly and functionally distinct subpopulations:
Calbindin-Positive vs. Calbindin-Negative Neurons: A subset of SNc neurons expresses calbindin-D28k, a calcium-binding protein that may confer neuroprotective properties. Calbindin-negative neurons in the ventrolateral tier show greater vulnerability in PD.
Aldehyde Dehydrogenase 1A1 (ALDH1A1): A specific marker identifies a more vulnerable SNc subpopulation. ALDH1A1-positive neurons are preferentially lost in PD and show increased susceptibility to oxidative stress.
FOXA2+ Neurons: The transcription factor FOXA2 defines a subset of SNc neurons critical for dopaminergic identity and survival. These neurons show early vulnerability in PD models.
¶ Pars Reticulata Organization
The SNr contains heterogeneous GABAergic neurons:
Type I vs. Type II Neurons: Two major populations can be distinguished electrophysiologically and neurochemically. Type I neurons project to the thalamus and superior colliculus, while Type II neurons primarily target the pedunculopontine nucleus.
Sensorimotor vs. Associative Territories: Functional mapping reveals somatotopic organization within the SNr, with distinct regions controlling different aspects of movement and cognition.
¶ Molecular Mechanisms of Vulnerability
¶ Calcium Dysregulation
SNc dopaminergic neurons exhibit unique electrophysiological properties that contribute to their vulnerability:
L-Type Calcium Channels: The Cav1.3 isoform generates autonomous pacemaking in SNc neurons. This requires continuous calcium influx, which stresses mitochondrial energy production.
Calcium-Binding Proteins: Unlike VTA neurons, SNc neurons have low calbindin content, limiting their ability to buffer calcium loads. This deficiency may contribute to selective vulnerability.
Calcium-Induced Mitochondrial Dysfunction: Chronic calcium influx promotes mitochondrial permeability transition, ROS generation, and ultimately cell death.
¶ Iron Homeostasis
The substantia nigra has the highest brain iron concentration:
DMT1 and Ferritin: Iron import via DMT1 and storage in ferritin are dysregulated in PD. Increased iron promotes Fenton chemistry and oxidative damage.
Ferroptosis: Iron-dependent lipid peroxidation may contribute to SNc degeneration. GPX4 activity and lipid metabolism are key regulators of this process.
Iron as a Biomarker: Quantitative susceptibility mapping (QSM) MRI reveals iron accumulation in the SNc of PD patients, correlating with disease severity.
¶ Mitochondrial Dynamics
SNc neurons show particular sensitivity to mitochondrial stress:
Complex I Deficiency: Rotenone and MPTP both inhibit complex I, reproducing PD pathology. Genetic forms of PD (PINK1, parkin) directly affect mitochondrial quality control.
Mitochondrial DNA Mutations: Accumulation of somatic mitochondrial DNA mutations in SNc neurons exceeds other brain regions, potentially due to high metabolic demand.
Mitochondrial Biogenesis: PGC-1α signaling, which drives mitochondrial biogenesis, is downregulated in PD. Enhancement of this pathway is protective in models.
¶ Synaptic Circuitry
¶ Nigrostriatal Pathway
The SNc to striatum projection is the core of motor control:
Terminal Organization: Single SNc neurons form 100,000-250,000 synaptic contacts in the striatum. Each neuron innervates ~6% of the striatal volume, providing widespread modulation.
Dopamine Release Patterns: Tonic dopamine release maintains baseline striatal activity, while phasic bursts encode reward prediction errors and salient events.
Striatal Microcircuitry: Dopamine modulates direct and indirect pathway medium spiny neurons differently, enabling fine motor control.
¶ Subthalamic Nucleus Interactions
The subthalamic nucleus provides critical excitatory input to the SNr:
Hyperdirect Pathway: Cortical input reaches SNr within 20 ms via the subthalamic nucleus, enabling rapid movement suppression.
Pathological Activity: In PD, excessive beta-frequency synchronization in the subthalamic nucleus drives excessive SNr output, producing bradykinesia and rigidity.
¶ Therapeutic Advances
¶ Gene Therapy Approaches
Several gene therapy strategies are in clinical development:
AADC Gene Therapy: AAV-mediated AADC delivery to the striatum enhances endogenous levodopa conversion. Promising results from trials show improved "on" time without dyskinesias.
GDNF and Neurturin: AAV-NRTN (CERE-120) showed initial promise but failed in phase 2 trials. Ongoing studies explore optimized delivery methods.
Alpha-Synuclein Silencing: siRNA and antisense oligonucleotides targeting SNCA mRNA reduce alpha-synuclein production in preclinical models.
¶ Cell Replacement Therapy
Stem cell-derived dopaminergic neurons offer potential for circuit restoration:
Embryonic Stem Cell-Derived Neurons: Clinical trials using ESC-derived midbrain dopamine progenitors (e.g., STEM-PD) are underway in Europe and Australia.
Induced Pluripotent Stem Cells: Patient-derived iPSCs enable personalized cell therapy and disease modeling. Several groups have initiated clinical-grade iPSC banking.
Immune Evasion: Gene editing to delete HLA molecules may enable universal donor cells without immunosuppression.
¶ Alpha-Synuclein-Targeting Immunotherapies
Active and passive immunization approaches target pathological alpha-synuclein:
Affitopes: AFFiRiS developed peptide-based vaccines (AFFITOPE PD01A) showing safety in phase 1. Antibodies against the C-terminus of alpha-synuclein were generated.
Passive Antibodies: Several monoclonal antibodies are in clinical trials:
- Cinpanemab (BIIB054): Targets aggregated alpha-synuclein
- Prasinezumab (PRX002): Shows slowing of motor progression in phase 2
Oligonucleotide Therapies: Antisense oligonucleotides targeting SNCA reduce protein production in model systems.
¶ Research Techniques
¶ Neuroimaging
- DaTSCAN (ioflupane SPECT): Visualizes dopamine transporter density in the striatum as a surrogate for SNc neuronal integrity
- Neuromelanin-sensitive MRI: Directly images neuromelanin-containing neurons in the substantia nigra, providing a non-invasive measure of SNc degeneration (Sulzer et al., 2018
- Iron-sensitive MRI (QSM): Quantifies iron accumulation in the substantia nigra
¶ Single-Cell Genomics
Recent single-cell-genomics studies have identified molecularly distinct dopaminergic subpopulations within the SNc. Kamath et al. (2022) identified a specific subpopulation expressing SOX6 and AGTR1 that is highly susceptible to neurodegeneration in PD and enriched for PD-associated GWAS risk genes (Kamath et al., 2022. Agarwal et al. (2024) further refined this [taxonomy, identifying ten transcriptomically distinct neuron types in the human substantia nigra (Agarwal et al., 2024.
- [Dopaminergic [Neurons (SNpc)/cell-types/dopaminergic-neurons-snpc
- alpha-synuclein
- deep-brain-stimulation
- levodopa
¶ External Links
- Substantia Nigra — StatPearls (NCBI)
- Substantia Nigra — Wikipedia
- Cleveland Clinic — Substantia Nigra
- Allen Human Brain Atlas — Substantia Nigra
¶ Background
The study of Substantia Nigra has evolved significantly over the past decades. Research in this area has revealed important insights into the underlying [mechanisms of neurodegeneration/mechanisms) and continues to drive therapeutic development.
Historical context and key discoveries in this field have shaped our current understanding and will continue to guide future research directions.
¶ Brain Atlas Resources
- Allen Brain Atlas
- Allen Human Brain Atlas
- Allen Mouse Brain Atlas
- Allen Cell Type Atlas
- BrainSpan Developmental Transcriptome
¶ Visual Resources
¶ Substantia Nigra Anatomical Schematic
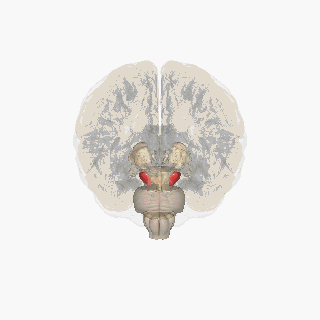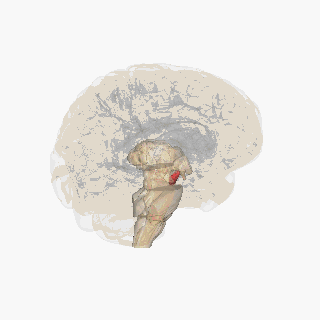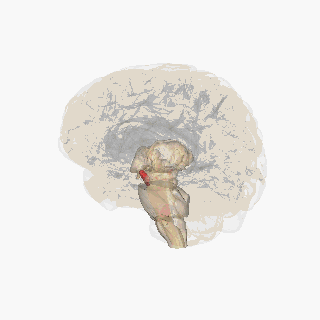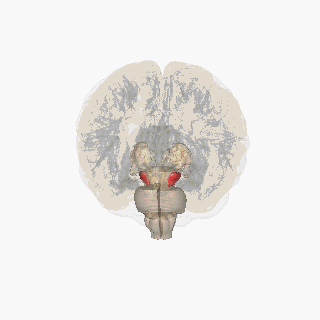
This schematic highlights the position and basic anatomy of the substantia-nigra, a central node in parkinsons pathophysiology.
Image attribution: FrozenMan, Substantia nigra (CC BY-SA 4.0)(https://commons.wikimedia.org/wiki/File:Substantia_nigra.gif)
{kind=link}
¶ Conclusion
The Substantia Nigra is a critical brain region in the pathophysiology of Parkinson's disease, containing the dopaminergic neurons of the SNpc that project to the striatum. The selective vulnerability of these neurons to [alpha-synuclein aggregation, mitochondrial dysfunction, and neuroinflammation makes the Substantia Nigra a central focus of PD research and therapeutic development. Understanding the molecular mechanisms underlying nigral degeneration, including Lewy body pathology spreading from the locus coeruleus, continues to inform disease-modifying strategies. Cell replacement approaches, including dopamine neuron transplantation and stem cell therapies, aim to restore nigrostriatal circuitry in PD patients.
¶ Research Evidence
¶ RNA sequencing and eQTL discovery in human putamen and substantia nigra
Identified 19,156 significant eQTL signals genome-wide at FDR 5%, including 359 secondary eQTLs. eQTL discovery was not simply driven by number of features tested; rate of eQTL discovery was highest in unannotated intronic and intergenic regions. 50.6% of putamen and 50.4% of substantia nigra eQTLs replicated in microarray data; 39.3% putamen and 50.6% substantia nigra eQTLs replicated in GTEx data.
Model System: Human postmortem brain tissue (putamen and substantia nigra from 117 neurologically healthy individuals of European descent)
Statistical Significance: FDR < 5%
Sebastian Guelfi et al., (2020)
¶ eQTL replication in external datasets
Replication rates: GTEx putamen 19-53.3%, GTEx substantia nigra 50.6-62.0%; PsychENCODE 64.2%; CommonMind 54.2%; Lappalainen lymphoblastoid cell lines only 22.0% putamen and 24.2% substantia nigra. Higher replication in brain tissues than lymphoblastoid cells.
Model System: Human brain datasets (GTEx, PsychENCODE, CommonMind) and lymphoblastoid cell lines
Statistical Significance: FDR < 5% in both datasets
Sebastian Guelfi et al., (2020)
¶ Cell type specificity analysis using weighted gene co-expression network analysis (WGCNA)
75% of analysed genes confidently assigned to specific cell type. eQTL target expression features assigned to cell types: 41.5%. Significant enrichment of neuronal genes in non-standard eQTL classes: i-eQTLs (FDR p=1.20x10^-2 in putamen), e-eQTLs (FDR p=1.21x10^-7 in substantia nigra), ex-ex-eQTLs (FDR p=2.28x10^-5 in substantia nigra). Splicing eQTLs also enriched for oligodendrocyte and astrocyte genes.
Model System: Human putamen and substantia nigra
Statistical Significance: FDR-corrected Fisher's Exact p-values: i-eQTLs p=1.20x10^-2, e-eQTLs p=1.21x10^-7, ex-ex-eQTLs p=2.28x10^-5
Sebastian Guelfi et al., (2020)
¶ Allele-specific expression (ASE) discovery and validation
Analysed 252,742 valid heterozygous SNPs. 19,266 (7.62%) were significant ASEs at FDR 5%, covering 8,654 genes. 12,096 ASEs in putamen, 11,871 in substantia nigra. 67% of testable ASE signals validated in lymphoblastoid cell lines. Inconsistent ASE signals (not unidirectional in >=10 individuals) were only in known imprinted genes (1.96% on X chromosome).
Model System: Human putamen and substantia nigra (subset with whole-exome sequencing)
Statistical Significance: FDR < 5%
Sebastian Guelfi et al., (2020)
¶ Cell type specificity of ASE signals
ASE-containing genes highly enriched for neuronally expressed genes: putamen FDR p=9.97x10^-235, substantia nigra FDR p=3.05x10^-97. Also significant enrichments for oligodendrocyte, astrocyte, microglia, and endothelial gene sets. Strength of evidence for cellular specificity was striking for ASEs compared to eQTLs.
Model System: Human putamen and substantia nigra
Statistical Significance: Putamen: FDR p=9.97x10^-235; substantia nigra: FDR p=3.05x10^-97
Sebastian Guelfi et al., (2020)
¶ Disease relevance of ASEs - GWAS enrichment
Highly significant enrichment of GWAS risk loci among ASEs: schizophrenia p=7.49x10^-35, Parkinson's disease p=4.19x10^-7. Using LD score regression, Parkinson's disease heritability enrichment appeared more specific to ASEs in substantia nigra. No enrichment in PD or schizophrenia heritability among eQTLs using same method.
Model System: ASE data overlapped with PD and schizophrenia GWAS
Statistical Significance: Schizophrenia: p=7.49x10^-35; Parkinson's disease: p=4.19x10^-7
Sebastian Guelfi et al., (2020)
¶ Dopaminergic neuron subtyping with classifier comparison to Kamath et al. dataset
Identified 14 TH+ subtypes (vs 10 in Kamath et al.). Found 7 new TH+ subtypes including SOX6 LPL subtype in substantia nigra and 4 CALB1+ subtypes in periaqueductal grey and VTA. Identified third class of TH+ neurons co-expressing GABAergic markers (CALCR and EBF2 clusters).
Model System: Human midbrain dopaminergic neurons from this study vs published dataset
Statistical Significance: Cross-validation performed with varying parameters
¶ Head-to-head comparison of tau PET tracers
Off-target binding profiles vary across tracers. 18F-flortaucipir shows off-target binding in basal ganglia, substantia nigra, longitudinal sinuses, pituitary, choroid plexus. 18F-RO948 and 18F-MK6240 show greater binding to meninges and skull. 18F-PI2620 shows promise for 4R tauopathies with lower off-target binding in basal ganglia.
Model System: Human participants
Statistical Significance: Not specified
¶ 18F-T807/AV-1451 evaluation in non-AD tauopathies
Regional tau PET patterns agreed with regions underlying clinical symptoms in atypical AD. FDG PET abnormally low where tau high. In PPA, tau signal spreading through language networks. In PSP, positive signals in substantia nigra, globus pallidus, and subthalamic nucleus, but some overlap with controls limiting early-stage detection.
Model System: PSP patients, primary progressive aphasia patients, atypical AD patients
Statistical Significance: p < 0.001 uncorrected for PSP vs controls in specific brain regions
Hartmuth C. Kolb, José Ignacio Andrés (2017)
¶ Neural Connectivity
¶ Comparative Anatomy
| Feature | Pars Compacta | Pars Reticularis |
|---|---|---|
| Primary Neurotransmitter | Dopamine | GABA |
| Primary Afferents | Striatum, Cortex | Striatum, Cortex |
| Primary Efferents | Striatum, Pallidum | Thalamus, Colliculi |
| Neurodegeneration in PD | Severe (60-80% loss) | Moderate |
| Key Cell Types | Dopaminergic neurons | GABAergic neurons |
¶ Disease Relevance
| Disease | SNc Involvement | Key Pathology |
|---|---|---|
| Parkinson's Disease | Severe neuronal loss | Lewy bodies (α-syn) |
| Progressive Supranuclear Palsy | Moderate | Tau pathology |
| Multiple System Atrophy | Moderate | Oligodendrocytic inclusions |
| Dementia with Lewy Bodies | Severe | Lewy bodies |
¶ Experimental Models and Research Tools
¶ Genetic Models
Alpha-Synuclein Models:
- SNCA transgenic mice: Overexpression of wild-type or mutant SNCA under various promoters produces Lewy-like pathology
- AAV-mediated overexpression: Direct injection of AAV-SNCA into rat SNc produces progressive degeneration
- Alpha-synuclein preformed fibrils: Inoculation with synthetic α-syn fibrils triggers pathological spreading
Parkinsonian Genetic Models:
- PINK1 knockout: Mitochondrial dysfunction and progressive dopamine neuron loss
- Parkin knockout: Subtle phenotypes, enhanced sensitivity to toxins
- LRRK2 G2019S knock-in: Late-onset progressive degeneration with minimal basal phenotype
¶ Toxin Models
- 6-Hydroxydopamine (6-OHDA): Retrograde degeneration of SNc neurons after striatal injection; complete unilateral lesions for lesion studies
- MPTP: Prodrug converted to MPP+ which inhibits complex I; produces acute parkinsonism in primates
- Rotenone: Chronic complex I inhibition; reproduces Lewy bodies in some models
- Paraquat: Increases oxidative stress; causes progressive SNc degeneration
¶ In Vitro Systems
- Dopaminergic cell lines: N27, MN9D cells for mechanistic studies
- Primary neuron cultures: Dissociated mesencephalic cultures
- Organotypic slice cultures: Maintain circuitry; enable long-term manipulation
- iPSC-derived neurons: Patient-specific disease modeling; disease-relevant phenotypes include:
- Reduced neurite complexity
- Increased α-synuclein aggregation
- Mitochondrial dysfunction
- Altered autophagic flux
¶ Advanced Research Techniques
Circuit Mapping:
- Retrograde tracing (CTB, rabies virus) for afferent mapping
- Anterograde tracing (AAV, PHAL) for efferent mapping
- Optogenetic mapping of functional connectivity
- Graph theory for network analysis
Electrophysiology:
- Patch-clamp recordings in vivo and in vitro
- Multi-electrode arrays for population activity
- Optogenetic manipulation with simultaneous recording
Molecular Profiling:
- Single-nucleus RNA sequencing
- ATAC-seq for chromatin accessibility
- Spatial transcriptomics (Visium, MERFISH)
- Proteomics of subcellular fractions
¶ Biomarkers for SNc Integrity
¶ Imaging Biomarkers
Dopamine Terminal Imaging:
- DaT-SPECT/Ioflupane: Measures dopamine transporter binding; sensitive to early degeneration
- FP-CIT PET: Alternative dopamine transporter ligand
- 123I-FP-CIT: FDA-approved for differentiating PD from non-parkinsonian tremor
Neuronal Integrity:
- Neuromelanin MRI: Direct visualization of SNc neurons; signal correlates with neuronal density
- Diffusion MRI: Elevated diffusion in degenerated SNc
- PET with receptor ligands: D2 receptor imaging as downstream marker
Iron and Metabolism:
- Quantitative Susceptibility Mapping (QSM): Elevated iron in SNc of PD patients
- R2 mapping*: Iron-sensitive MRI technique
- FDG-PET: Shows characteristic pattern of hypometabolism in basal ganglia
¶ Fluid Biomarkers
Neurodegeneration Markers:
- Neurofilament light chain (NfL) in CSF and blood — correlates with disease progression
- Total tau and phosphorylated tau — altered in some PD patients
α-Synuclein:
- Seed amplification assay (RT-QuIC, PMCA) detects pathological α-synuclein
- CSF α-synuclein: reduced total, increased phosphorylated
- Blood extracellular vesicles — promising for peripheral detection
Inflammation:
- CSF cytokine profiles (IL-6, TNF-α)
- Monocyte activation markers
¶ Conclusion
The substantia nigra, particularly the pars compacta, stands at the center of Parkinson's disease pathophysiology. Its selective vulnerability stems from the convergence of multiple factors: high metabolic demand from extensive axonal arbors, autonomous pacemaking driven by calcium influx, dopamine metabolism generating reactive oxygen species, and the age-related accumulation of neuromelanin-bound iron.
Understanding the molecular mechanisms underlying SNc neurodegeneration has yielded promising therapeutic strategies. From dopamine replacement with levodopa to advanced gene therapies, cell replacement approaches, and alpha-synuclein-targeted immunotherapies, the therapeutic landscape continues to evolve. The identification of molecularly distinct vulnerable subpopulations through single-cell genomics offers hope for more targeted interventions.
As biomarker development progresses, earlier detection becomes possible — potentially enabling disease-modifying treatments before irreversible neuronal loss occurs. The continued integration of experimental models, human tissue studies, and clinical research promises to accelerate translation from basic discovery to effective therapies.
¶ References
- Zecca L, Zucca FA, Albertini A, Rizzio E, Fariello RG, A proposed dual role of neuromelanin in the pathogenesis of Parkinson's Disease (2006)
- Damier P, Hirsch EC, Agid Y, Graybiel AM, The substantia nigra of the human brain (1999)
- Lanciego JL, Luquin N, Obeso JA, Functional neuroanatomy of the basal ganglia (2012)
- Björklund A, Dunnett SB, Dopamine neuron systems in the brain: an update (2007)
- Pakkenberg B, Moller A, Gundersen HJ, Mouritzen Dam A, Pakkenberg H, The absolute number of nerve cells in substantia nigra in normal subjects and in patients with Parkinson's Disease estimated with an unbiased stereological method (1991)
- Matsuda W, Furuta T, Nakamura KC, et al, Single nigrostriatal dopaminergic neurons form widely spread and highly dense axonal arborizations in the neostriatum (2009)
- Deniau JM, Mailly P, Maurice N, Charpier S, The pars reticulata of the substantia nigra: a window to basal ganglia output (2007)
- Gerfen CR, Surmeier DJ, Modulation of striatal projection systems by dopamine (2011)
- Zhang W, Phillips K, Wielgus AR, et al, Neuromelanin activates microglia and induces degeneration of dopaminergic neurons: implications for progression of Parkinson's Disease (2011)
- Kalia LV, Lang AE, Parkinson's Disease (2015)
- Spillantini MG, Schmidt ML, Lee VM, Trojanowski JQ, Jakes R, Goedert M, alpha-synuclein in Lewy bodies (1997)
- Braak H, Del Tredici K, Rüb U, de Vos RA, Jansen Steur EN, Braak E, Staging of brain pathology related to sporadic Parkinson's Disease (2003)
- Surmeier DJ, Obeso JA, Halliday GM, Selective neuronal vulnerability in Parkinson disease (2017)
- Pacelli C, Giguère N, Bhose MJ, et al, Elevated mitochondrial bioenergetics and axonal arborization size are key contributors to the vulnerability of dopamine neurons (2015)
- Chan CS, Guzman JN, Ilijic E, et al, 'Rejuvenation' protects neurons in mouse models of Parkinson's Disease (2007)
- Dexter DT, Wells FR, Lees AJ, et al, Increased nigral iron content and alterations in other metal ions occurring in brain in Parkinson's Disease (1989)
- McGeer PL, Itagaki S, Boyes BE, McGeer EG, Reactive microglia are positive for HLA-DR in the substantia nigra of Parkinson's and Alzheimer's Disease brains (1988)
PMID:19296920 - Booth HDE, Hirst WD, Wade-Martins R, The role of astrocyte dysfunction in Parkinson's Disease pathogenesis (2017)
PMID:29203312 - Limousin P, Krack P, Pollak P, et al, Electrical stimulation of the subthalamic nucleus in advanced Parkinson's Disease (1998)
- Kamath T, Abdulraouf A, Burber SJ, et al, Single-cell genomic profiling of human dopamine neurons identifies a population that selectively degenerates in Parkinson's Disease (2022)
- Agarwal D, Sandor C, Volpato V, et al, Molecular profiling of human substantia nigra identifies diverse neuron types associated with vulnerability in Parkinson's Disease (2024)